

В развитии животных имеется особый период зародышевого сходства, во время которого не только облик эмбрионов, но и экспрессия их генов весьма консервативны. Существование этого периода связано с появлением в эволюции устойчивых планов строения. Как и почему сформировался этот консервативный период эмбриогенеза? Исследование линий нематоды C. elegans с повышенным числом мутаций показало, что мутации мало затрагивают этот консервативный этап развития, сильнее срабатывая в начале и в конце развития. На этом этапе наиболее активно работают HOX-гены и гены, координирующие развитие зародышевых листков. По мнению авторов, именно эта координация является главным фактором, ограничивающим изменчивость эмбрионального развития.

XXI век заставил ученых постепенно отказаться от жестких установок биогенетического закона Геккеля-Миллера о повторении филогенеза в онтогенезе. Закон, напомню, основан на видимом сходстве эмбрионов того или иного организма со взрослыми формами, причем последовательный ряд эмбриональных портретов повторяет череду эволюционных этапов, которые прошел данный организм. Сейчас ученые пытаются говорить о сходстве эмбриональных серий, а не взрослых портретов, о сходстве только некоторых из эмбриональных стадий, или же вообще предлагают отказаться от этого закона.
Но в случае полного отрицания биогенетического закона останется необъяснимой исключительная консервативность планов строения в пределах каждого типа (собственно, таксономические типы животных группируются на основе сходства их плана строения). Ведь закон Геккеля выполнял функцию логичного истолкования этой консервативности: раз имеется эволюционная преемственность морфологий в ходе эмбриогенеза, то и план строения, размеченный в ходе эмбрионального развития, должен сохраняться.
Но в арсенале биологов есть еще один закон, который никто не отменял, — закон Бэра о зародышевом сходстве. Этот закон (или, скорее, набор правил) утверждает, что на определенной эмбриональной стадии зародыши у всех представителей типа похожи, а затем постепенно обретают свои специфические черты. Теоретическое осмысление этих правил и их мощная фактологическая поддержка привели к описанию так называемых филотипических стадий эмбрионального развития (термин предложен в 1983 году Клаусом Сандером). «Фило-» в данном случае относится не столько к эволюции, сколько к таксономии, обозначая термин «тип животного» (греч. phylon).
Иначе говоря, филотипическая стадия — это особый этап эмбриогенеза, на котором эмбрионы в пределах каждого плана строения (типа) обнаруживают высокое сходство друг с другом. Раньше и позже филотипической стадии эмбрионы похожи друг на друга гораздо меньше. Именно на этой стадии в основном и определяется план строения животного.
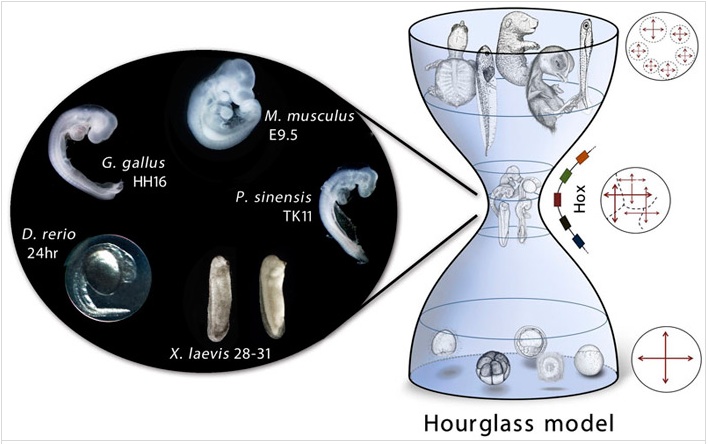
Наличие филотипической стадии сейчас подтверждено для нескольких типов животных (насекомые, нематоды, моллюски, кольчатые черви, позвоночные). Есть, правда, и исследования, в которых существование такой консервативной стадии подвергается серьезному сомнению. Кроме того, предложены и другие варианты для описания консервативного зародышевого сходства (см. статью Е. Северцова, А. Северцов, 2013. Критические периоды в постэмбриональном развитии остромордой лягушки (R. arvalis) и ссылки в ней). Но, так или иначе, наличие «узкого места» в череде зародышевых портретов нужно считать установленным фактом. Его изображают в виде песочных часов, указывая на снижение вариабельности зародышевых фенотипов в определенный период эмбриогенеза.
Последние исследования показывают, что в этот период не только фенотипы, но и экспрессия генов сильно зарегулирована. Разброс в уровне экспрессии генов оказывается минимален во время филотипического периода в отличие от более поздних и более ранних этапов. То есть картина экспрессии генов тоже повторяет абрис песочных часов, хотя жесткой синхронности пережимов у «феночасов» и «геночасов» нет. Нужно отметить, что подобные песочные часы на уровне экспрессии генов были определены и для растений (см.: М. Гельфанд, Песочные часы в биологии развития), и даже для грибов. Сейчас, правда, для арабидопсиса, единственного пока растения, для которого проверили наличие филотипического этапа эмбриогенеза, выявили не один, а несколько таких «узких» мест — на стадии прорастания и цветения (H.-G. Drost et al., 2016. Post-embryonic Hourglass Patterns Mark Ontogenetic Transitions in Plant Development). Так что сравнивать филотипические стадии животных и растений (и, тем более, грибов) пока рановато.
Какие гены работают на филотопической стадии эмбриогенеза? Принципиально возможны два пути для поиска ответа на этот вопрос. Первый открывает сравнительная транскриптомика. Она предлагает сравнивать уровень транскрипции генов-ортологов у видов, разделенных разной степенью родства. Второй путь — филотранскриптомика. Этот подход возможен, если имеются данные по уровню транскрипции широкого набора генов у одного вида. И сравнительная транскриптомика, и филотранскриптомика показывают, что на уровне пережима песочных часов активно экспрессируются HOX-гены, которые размечают план строения животного.
В отличие от закона Геккеля, надежно увязанного с эволюционной историей, филотипическую стадию пока трудно сопоставить с эволюционным развитием групп или хотя бы с работой отбора. Предполагается, что узкое место в эмбриогенезе может появляться вследствие действия стабилизирующего отбора, который отбрасывает все варианты, отклоняющиеся на этой особой стадии развития. Таким образом у песочных часов может образоваться талия. Но вероятен и другой механизм: движущий отбор сильнее действует на ранние и поздние стадии эмбриогенеза, так как для адаптации организма они важнее. И таким образом «верх» и «низ» песочных часов расширяются. А может быть и так, что стабилизирующий и движущий отборы действуют совместно.
В интересном эксперименте эмбриологи Арель Зальц (Harel Zalts) и Итаи Янаи (Itai Yanai) рассмотрели, что произойдет с филотипической стадией, если вообще снять давление отбора. Сохранится ли она или нет? А если сохранится, то какие гены к ней привязаны больше всего?
Ученые работали c нематодами Caenorhabditis elegans. Благодаря очень короткому жизненному циклу этих нематод (3 дня), эксперимент занял «всего» два года. В течение этого времени ученые выводили от одного общего родителя 19 линий, в которых отбор полностью отсутствовал и шло накопление мутаций. Для этого для каждого следующего поколения случайным образом выбирался один гермафродитный основатель, который давал потомков, и раунд со случайным выбором основателя повторялся. И так — 250 раз. Ясно, что мутации — и вредные, и нейтральные, и полезные — могли появляться и накапливаться в каждой линии случайным образом.
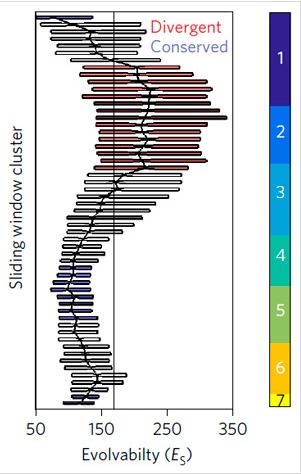
Через два года в полученных линиях был проверен уровень экспрессии генов, работающих на разных этапах зародышевого развития. Развитие C. elegans принято подразделять на 7 стадий. Консервативной, то есть филотипической, считается четвертая стадия (стадия брюшного замыкания). Итак, осталась ли после накопления мутаций эта стадия столь же консервативной?
Осталась. По крайней мере, на уровне транскриптома. Подсчитав изменчивость наступления и интенсивности экспрессии (то есть оценку гетерохронии в работе каждого гена), ученые нарисовали график этой изменчивости. Он имеет явный минимум на уровне 4-5 стадий зародышевого развития.
Наиболее активные в этот период гены относятся к двум функциональным группам. Это гомеобоксные гены и гены, которые отвечают за координированное развитие зародышевых листков. Это те гены, которые не экспрессируются, если клетки энтодермы, эктодермы и мезодермы культивировать отдельно. Их работа выявляется только при работе с реальными эмбрионами. Авторы назвали их генами интеграции и подчеркнули, что именно эта группа генов оказалась наиболее консервативна в их эволюционном эксперименте. Даже больше, чем HOX-гены.
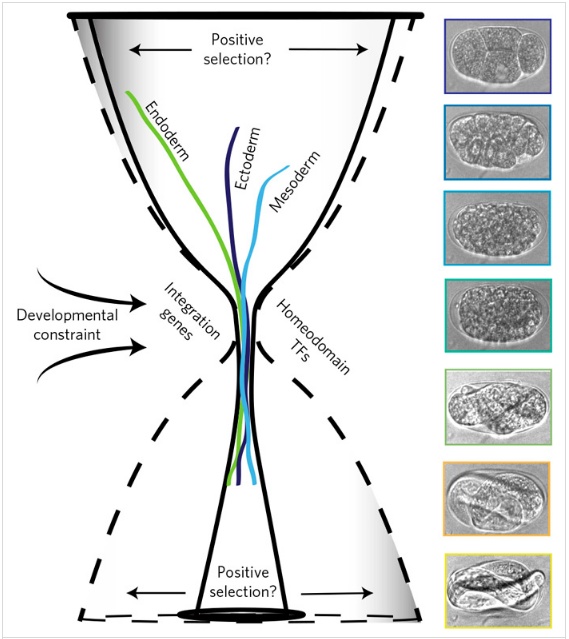
Обсуждая результаты, ученые делают важное замечание относительно сужения диапазона изменчивости на последних стадиях развития (6-й и 7-й). Если сравнивать филогенетически далекие виды того или иного типа (результаты сравнительной транскриптомики), то в песочных часах более широкой окажется верхняя (поздняя) часть, а если раснивать близкие виды или изменчивость в пределах одного вида (филотранскриптомика), то более широкой будет нижняя (ранняя) часть. Этот эффект отражает, по-видимому, сильную зарегулированность поздних стадий развития у близких видов. Ведь именно они предшествуют появлению организма, который обязан вести борьбу за существование и оставить потомство, то есть — проявить финальную приспособленность. В представленном примере с филотранскриптомным анализом С. elegans наблюдается, по всей вероятности, именно этот эффект.
Результаты работы позволяют заключить, что кратковременное снятие давления отбора не снимает ограничений с процесса развития. Он все равно остается зарегулированным в период становления плана строения, во время филотипической стадии. Существуют сильнодействующие внутренние факторы, определяющие ход развития. Чтобы ослабить их действие, нужны более серьезное влияние, чем 250 поколений случайного накопления мутаций.
В теоретическом смысле интересна (и достойна дальнейшего серьезного обсуждения) авторская гипотеза о причинах появления филотипических стадий. Она явно возникла из нового факта, который обнаружился в ходе этого исследования: высокой консервативности интеграционных генов. Согласно этой гипотезе, консервативный период должен был сформироваться при появлении двуслойности у животных. На ранних этапах развития экспрессируются гены энтодермы, она имеет более древний возраст и соответствует гипотетическим однослойным животным. Затем появилась эктодерма и, соответственно, двуслойные животные. Филотипический период обозначает переход от однослойности к двуслойности, когда потребовалось включить новые генетические приемы, увязывающие развитие двух слоев. Так что, по мнению авторов, консервативный период в эмбриогенезе — это наследие регуляторных сложностей, с которыми столкнулись и которые преодолели генетические системы животных. Добавление третьего зародышевого листка еще более усложнило регуляторную задачу.
Автор: Елена Наймарк. Источник: Elementy.ru
Дополнительно:
«Истина существует, и целью науки является ее поиск»
По следам этих невероятных человеческих следов.
Свидетельства и доказательства эволюции.
Был ли Гитлер атеистом, а его режим - атеистическим.
Первая любовь Чарльза Дарвина.
Эмбрионы Геккеля: мошенничество не доказано.
Урок биологии №11. Виды размножения.
Урок биологии №20. Грегор Мендель и его Первый закон.
Урок биологии №41. Эмбриональное развитие.
Мейоз. Фазы мейоза. Урок биологии №57
Урок биологии №51. Методы биологических исследований.




Оценили 17 человек
29 кармы